El citoplasma bacterià es la masa de materia viva delimitada per una membrana citoplasmàtica, al interior del qual trobem: cosos nuclears (nucleòide), plasmidis (no en totes les cepes bacterianes), ribosomes, inclusions i orgànuls.
Dos d’aquestes formacions, les vesícules i les inclusions citoplasmàtiques, són vitals en la supervivència i l’adaptació de molts tipus de bacteris.
Una vesícula és un orgànul fonamental, present en moltes cèl·lules, per mantenir l’organització del metabolisme. Forma un compartiment petit i tancat separat del citoplasma per una bicapa lipídica. La seva funció és emmagatzemar, transportar i digerir els productes i residus cel·lulars. Moltes d’aquestes són formades a l’aparell de Golgi (AG), però també en el reticle endoplasmàtic o a partir de parts de membrana plasmàtica.
Degut a la seva simplicitat morfològica (en comparació amb organismes eucariotes que tenen més facilitats d’adaptació), els bacteris han desenvolupat formes d’adaptació en forma d’orgànuls o inclusions que els permeten un major rang de supervivència depenent de les condicions ambientals. Així doncs augmenten les seves possibilitats de supervivència en diferents hàbitats.
1. INCLUSIONS CITOPLASMÀTIQUES
Les inclusions són acumulacions de substàncies orgàniques (fonts de carboni o nitrogen) o inorgàniques (fonts de fòsfor o sofre) que s’originen dins el citoplasma en determinades condicions de creixement. Poden estar o no limitades per una membrana de naturalesa proteica. Aquestes, emmagatzemen nutrients, productes d’excreció i grànuls de pigments.
A continuació s’exposen alguns dels tipus d’inclusions i vesícules que podem trobar en microorganismes, i la seva utilitat. Dins de les inclusions, hi trobem les inclusions inorgàniques i les orgàniques. Aquestes les separem segons si el que sigui utilitzat;
Inclusions orgàniques
- Inclusions polisacàrides
Són acumulacions de glucans amb ramificacions, principalment midó i glicogen, que es dipositen de manera uniforme per tot el citoplasma quan determinats bacteris creixen en mitjans amb limitació de font de nitrogen, però on encara siguin abundants les fonts de carboni i energia. Quan les cèl·lules disposen d'un medi ric en nitrogen, però sense font de carboni, aquestes inclusions s'utilitzen com a font interna de carboni per a la síntesi d'àcids nucleics i proteïnes.
Per observar-les s'utilitza la tinció de iode present als grànuls de glucògen els quals es tornen de color vermell i els grànuls de midó que es tornen de color blau.
- Grànuls de Poli-ß-hidroxibutíric (PHB)
Els grànuls de poli-b-hidroxibutíric són agrupacions de polièster de l'àcid ß-hidroxibutíric (3-hidroxibutíric), envoltats d'una capa proteica, i igual que en el cas anterior, es produeixen en certs bacteris com a reserva osmòticament inert de carboni en condicions de falta de nitrogen.
En les espècies de Bacillus constitueix la font de carboni i energia a l'inici de l'esporulació. La síntesi es produeix per una branca lateral de la ruta de síntesi dels àcids grassos, mitjançant ß-hidroxibutiril-CoA. En els grànuls, el polímer queda associat a un complex que s'utilitzarà en la degradació, però aquest necessita una activació prèvia.
A diferència de les agrupacions de polisacàrids, els grànuls de PHB són visibles amb un microscopi òptic i en fresc, degut al seu elevat índex de refringència, es tenyeixen amb negre sudan.
- Grànuls de poli-ß-hidroxialcanoats (PHA):
Els grànuls de PHB són un exemple d’una classe més ample de grànuls de poli-ß-hidroxi-alcanoats. Algunes soques de Alcaligenes eutrophus, quan creixen en glucosa y propiònic produeixen copolímers d'unitats de ß-hidroxibutíric y ß-hidroxivalèric (3-hidroxipentanoic).
Els PHA es comporten com a excel·lents termoplàstics biodegradables. Per exemple, l'empresa britànica ICI té patentats processos industrials per fabricar PHA on quasi el 90% de les unitats són d'hidroxivalèric, que dóna un polímer flexible comercialitzat amb el nom de Biopol. Els polímers a base de 4- o 5-hidroxibutíric i 3-hidroxibutíric són més llargs, més elàstics i més biodegradables.
- Grànuls de cianoficina
Els grànuls de cianoficina són inclusions de reserva que apareixen com a grans grànuls de lipoproteïnes refrigerant de reserves nitrogenades al citoplasma de les cianobacteries sense membrana. Es creu que intervenen en la gènesis de la ficocianina, que és el pigment blau lliure de metalls en proteïnes conjugades d’algues blau-verdoses. El número d’aquests grànuls incrementa quan el medi és pobre en fosfats.
Les cianobacteries acumulen aquest material quan estan a la fase estacionària però si hi ha canvi de medi, el creixement es reinicia amb la degradació d'aquests grànuls. El pèptid dels grànuls de cianoficina és l'únic material de reserva nitrogenat orgànic present en procariotes.
- Carboxisomes
Els carboxisomes són inclusions que contenen l'enzim ribulosa 1,5-difosfat carboxilasa (RUBISCO). Com que els organismes fotosintètics només tenen com a font de carboni el diòxid de carboni necessiten aquest enzim el qual la seva funció principal és fixar diòxid de carboni. Entre els bacteris més important que contenen carboxisomes trobem els bacteris nitrificants, els cianobacteris i els tiobacils.
Inclusions inorgàniques
- Grànuls de polifosfats
Aquests grànuls son acumulacions de polifosfats, polímers lineals d’ortofosfats, que són els fosfats més avançats els quals contenen l’anió PO43-. Per als microorganismes representen una manera eficaç d’emmagatzematge de fosfat. Els grànuls de polifosfats tenen una gran utilitat sobretot en l’eliminació de fosfats en aigües residuals. Un dels bacteris que més profit en treu es Acinetobacter el qual pot arribar a acumular el 24% de la seva biomassa sota la forma de polifosfats.
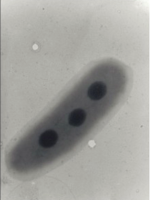 |
Imatge 1: grànuls de polifosfats en un bacterià del gènere Pseudomonas
|
- Glòbuls de sofre
Aquets glòbuls son característics de bacteris que utilitzen el sofre com a font de carboni en forma de SH2. Hi ha dos grups de bacteris:
Els bacteris porprats del sofre: són bacteris fotòtrofs on el donador d’electrons és el Sofre. Els pigments de bacterioclorofil·la a o b juntament amb carotenoides son els que els hi proporciona colors com el porpra, vermell i marró.
Bacteris filamentosos no fotosintètics com Beggiatoa o Thiothrix.
En tots dos casos el sofre elemental S0 de l’oxidació de sulfit s’acumula en aquests grànuls visibles. El sulfur romandrà dins el bacteri sempre que la font de sofre sigui present. Si la font de sulfur disminuís i es tornés limitant, el sulfur seria oxidat a sulfat (SO42-), i els grànuls anirien desapareixent poc a poc.
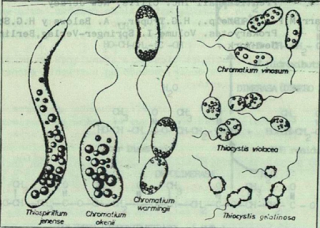 |
Imatge 2: diferents bacteris amb glòbuls de sofre
|
- Magnetosomes
Els magnetosomes són inclusions d'òxid de ferro produides per varis bacteris de la familia de gramnegatius com Magnetosperrilhon magnetotacticum. Aquests orgànuls intercel·lulars contenen cristalls de magnetita (Fe3O4) que els permet orientar-se cap a la Terra pel magnetisme. Els bacteris utilitzen aquestes inclusions per desplaçar-se per exemple per trobar un espai de fixació més apropiat.
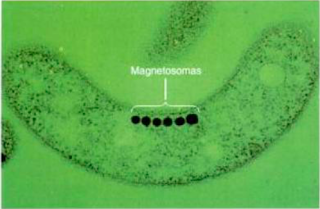
Imatge 3: Magnetosoma de Magnetospirillum magnetobacticum
2. VESÍCULES
Les vesícules són petits compartiments membranosos que viatgen entre orgànuls cel·lulars, i també a la membrana cel·lular. Serveixen per transportar molècules solubles, dissoltes en el medi aquós del seu interior, i molècules de membrana, que viatgen formant part de la pròpia membrana de la vesícula, com són lípids, canals o receptors. Una vegada les vesícules s’han format i carregat amb aquelles molècules que han de transportar, són alliberades al citosol i dirigides fins l’orgànul diana, al qual reconeixen, i es fusionen. Aleshores, les molècules transportades formaran part de l’orgànul diana i seran les responsables de la seva funció.
Tot i això, altres molècules només estaran de pas en aquest compartiment i seran empaquetades de nou en altres vesícules per dirigir-se a un altre compartiment cel·lular. Per exemple, algunes proteïnes viatgen des del reticle endoplasmàtic, passen per l’aparell de Golgi, on són empaquetades cap a altres compartiments. Altres tornaran a l’orgànul inicial a través d’una via de reciclatge.
Existeixen diferents tipus de vesícules: les vacuoles, els lisosomes i les vesícules de transport. Altres tipus de vesícules són les vesícules de gas utilitzades pels Archaea, bacteris i altres organismes planctònics. També existeixen vesícules extracel·lulars que serveixen per la biomineralització de teixits com ossos o cartílags.
Tot i això, altres molècules només estaran de pas en aquest compartiment i seran empaquetades de nou en altres vesícules per dirigir-se a un altre compartiment cel·lular. Per exemple, algunes proteïnes viatgen des del reticle endoplasmàtic, passen per l’aparell de Golgi, on són empaquetades cap a altres compartiments. Altres tornaran a l’orgànul inicial a través d’una via de reciclatge.
Existeixen diferents tipus de vesícules: les vacuoles, els lisosomes i les vesícules de transport. Altres tipus de vesícules són les vesícules de gas utilitzades pels Archaea, bacteris i altres organismes planctònics. També existeixen vesícules extracel·lulars que serveixen per la biomineralització de teixits com ossos o cartílags.
Vesícules de gas
Són estructures proteiques essencialment rígides per suportar la pressió. És hidrofòbica a l’aigua, però permeable als gasos, fet que fa que la composició i concentració del gas dins la vesícula depengui de les que existeixin en el medi. A mesura que es sintetitzen les vesícules, l’aigua va sent eliminada a l’interior. La seva morfologia i el nombre de vesícules varia segons l’organisme. Són presents en bacteris heteròtrofs, fotosintètics i fins i tot en alguns Archea. Aquestes vesícules, al estar carregades de gas consegueixen baixar la densitat de la cèl·lula, per tant aquesta s’eleva. En canvi, si les vesícules pujen, la cèl·lula baixa. Les vesícules de gas estan constituïdes per dos tipus de proteïnes: la majoritària s’anomena GvpA i en menor quantitat la GvpC.La GvpA és una petita proteïna molt rígida i hidrofòbica. La seva rigidesa és deguda a les pressions externes. D’altra banda, la GvpC té com a funció reforçar les vesícules de gas.A grans trets, les seves utilitats són la permeabilitat selectiva, possibilitat de flotació (augmentant o disminuint la densitat de la cèl·lula: poden regular a la fondària a la que es troba l’organisme en la columna d’aigua) i rigidesa (resistència a pressions externes).

Imatge 4: Vesícules de gas
CONCLUSIONS
En conclusió podem afirmar que molts microorganismes han desenvolupat adaptacions a medis i fonts per produir energia. Aquestes inclusions i vesicules desenvolupades donen pas a l'evolució de les espècies. No només per la utilitat que els hi ofereix als microorganismes la capacitat de poder utilitzar aquestes inclusions i vesícules sinó l’aportació que tenen per a altres ecosistemes incloent-hi el nostre.
BIBLIOGRAFIA I WEBGRAFIA
1. Megías Pachecho, M. (2015). Vesículas recuperat de http://mmegias.webs.uvigo.es/5-celulas/ampliaciones/5-vesiculas.php
2. Gerard J. Tortora, Berdell R. Funke, Christine L. Case. (2007). Introducción a la microbiología. Bases de la microbiología. 4: 96-97.
3. Ryan, V (2014). Biopol - Biodegradable Plastic. Recuperat de http://www.technologystudent.com/prddes1/biopola.html
Gotas de azufre. Recuperat de http://cdigital.dgb.uanl.mx/la/1020111466/1020111466_024.pdf
4. Madigan, M., Martinko, J., Parker, J. (2003). Brock. Biologia de los microorganismos. (10a ed.). Pearson Education.
5. Iáñez, E. (1998). Citoplasma. Inclusiones citoplasmáticas recuperat de http://www.biologia.edu.ar/microgeneral/micro-ianez/08_micro.htm
ANNEX
Imatge 1: Albieni, J. (2011). La célula. Como sé estructura, como se divide y se diferencia. Recuperat de http://jorgealbieni.blogspot.com.es/2011_05_01_archive.html
Imatge 2: Francisco P. Chávez Ph.D. Polifosfatos. Recuperat de http://www.bioblogia.com/polifosfatos/
Imatge 3: Gotas de azufre. Recuperat de http://cdigital.dgb.uanl.mx/la/1020111466/1020111466_024.pdf
Imatge 4: Gerard J. Tortora, Berdell R. Funke, Christine L. Case. (2007). Introducción a la microbiología. Bases de la microbiología. 4: pàg 96.
Cristina Badosa - Júlia Carbó - Judit Fernández - Andrea Garcia
Treball purament descriptiu en forma de llistat. Falta una mica d'estructura.
ResponEliminaPer què il.lustres amb una cèl.lula eucariota? Se suposa que parlem de bacteris.
Les inclusions no són vesícules, però no queda prou clar en el text. De fet les vesícules no hi apareixen més enllà de les "vesícules d'excreció" que cites (donant-li el rang d'apartat amb títol i tot) i que sembla que formen part de les cèl.lules de mamífers !!!!!????
No sé si vas venir a classe, però vam parlar de les vesícules de gas, que són un bon exemple.
Cal organitzar millor el treball.
primer distingir les vesícules de les inclusions oferint una definició vàlida d'ambdúes i, segon, organitzant millor els capítols. Un exemple: per què poseu els carboxisomes i magnetosomes apart? Que no són inclusions orgàniques i inorgàniques, respectivament?
Pel que va al desenvolupament, tt quedes en una descripció potser massa bàsica. Podries aprofitar els caràcters que tens per aprofundir una mica mes. També podries aprofitar i explicar millor què és això de les "membranes proteiques".... (Pista: GVP)
Faltaria veure en una taula comparativa quina és la distribució dels diferents elements descrits en el món microbià citant a quins grups pertanyen, si són aquàtics o no.
Editar millor la biblio (web)grafia...
No s'enten molt bé què es vol dir a la conclusió.
PREGUNTA EVALUABLE
ResponElimina¿Una sola bacteria puede tener inclusiones orgánicas e inorgánicas? ¿O el hecho de tener inclusiones de un tipo imposibilita tener del otro?
PABLO FERNÁNDEZ FERNÁNDEZ
Hola Pablo!
EliminaHem estat buscant informació sobre la teva pregunta i no hem trobat cap exemple amb el qual puguem justificar la teva pregunta. Tot i així, creiem que si un bacteri presenta inclusions orgàniques, és a dir, acumulacions de substàncies orgàniques (fonts de carboni o nitrogen) no tindrà inclusions inorgàniques ja que no utilitzarà el fòsfor o el sofre com a fonts d'energia per crèixer.
COMENTARI AVALUABLE:
ResponEliminaHola!
Pel que fa a la següent frase de la conclusió: “[...] sinó l’aportació que tenen per a altres ecosistemes incloent-hi el nostre”, em podríeu posar un exemple del que poden aportar aquests bacteris?
Moltes gràcies!
NOELIA GÓMEZ AGUILERA
Hola Noelia,
EliminaUn exemple molt fàcil d'explicar serien les vesícules de gas les quals estan presents en alguns bacteris com els cianobacteris i compleixen amb la funció de moviment en organismes que no disposen de flagels o altres sistemes de moviment. Així doncs permeten als organismes que les contenen flotar o enfonsar-se depenent de les necessitats o dels factors ambientals.
Els cianobacteris popularment coneguts com “algues blauverdes” són capaços de realitzar la fotosíntesi, i figuren entre les formes més antigues de la Terra. Porten al nostre planeta uns 3.500 milions d’anys, des d’una època tan diferent a la actual que en aquell moment les condicions eren anaeròbiques. De fet, per el que es sap, van ser els cianobacteris els que van oxigenar el planeta, instaurant les condicions que van permetre a la vida evolucionar cap a formes més complexes.
Avui en dia els cianobacteris encara formen una gran part dels nostres ecosistemes com per exemple els cianobacteris que es troben flotant als llacs, gràcies a l’acció de les vesícules de gas. Un exemple evident és el llac d’Atatlan a Guatemala que presenta fluorescència per causa dels cianobacteris.
Esperem haver resolt els teus dubtes!
Mitjançant quin procés els bacteris, com ara el esmentat (Acinetobacter), poden acumular polifosfats i formar així aquest tipus d'inclusió inorgànica?
ResponEliminaGràcies. Viviana Castro Martínez
Hola Viviana!
EliminaEn la degradació de polifosfats hi han tres possibles rutes. Per això, la seva inhibició es dóna en situacions d’alta relació ATP/ADP, i això es produeix en presència d’acceptors d’electrons, principalment oxigen. En resum, l’acumulació de polifosfats a l’interior de les cèl·lules s’afavoreix en condicions aeròbies mentre que l’absència d’oxigen afavoreix la hidròlisi.
En aquesta fase aeròbia, es disposa d’oxigen com acceptor d’electrons. La quantitat de substrat degradable en el medi és molt baix, tot i que els microorganisme posseeixen material orgànic de reserva (PHB) en el seu interior. La presència de O2 fa disminuir la relació NADH/NAD+ i estimula el cicle ATC i el del glioxilat, amb el que aconsegueix la oxidació del PHB i la obtenció de carboni per la síntesi cel·lular i d’energia. D’altra banda, l’oxigen permet la formació d’ATP i estimula la síntesi de polifosfat.
La quantitat de fòsfor que s’elimina es major que la que es redissol a la fase anaeròbia, aleshores es produeix un creixement cel·lular a partir de les reserves orgàniques i la energia generada en aquesta fase.
De totes les bacteries acumuladores de poifosfat, les més nombroses són Acinetobacter, que poden representar el 80 - 90% del total de microorganismes del fang.
Espero haver ressolt els teus dubtes!
L'autor ha eliminat aquest comentari.
ResponEliminaComentari avaluable:
ResponElimina-Les inclusions citoplasmàtiques apart de la possible capa proteica tenen alguna altre membrana que les envolti? si no es així són simples agregats de molècules? hi ha algun agent agregant o algun organul que les formi? i com us ha comentat el profe, quines són les diferencies essencials per diferencià una vesícula i una inclusió?
Alexis Salinas Alarcón
Bon dia Alexis!
EliminaLes inclusions són substàncies mortes que no són capaces de portar a terme qualsevol activitat metabòlica i no estan vinculades per membrana. Les inclusions són nutrients emmagatzemats, productes de secreció i grànuls de pigmentació.
En canvi, les vesícules emmagatzema, transporten o digereixen productes i residus cel·lulars.
Per tant, la major diferencia seria que les inclusions no tenen activitat metabòlica mentre que les vesícules són una eina essencial de la cèl·lula per la organització del metabolisme.
Espero que et serveixi d'ajuda!
Great post! I’ve been reading more about the benefits of NAD+ injections , and it’s amazing how they can boost energy, improve mental clarity, and support overall wellness. It’s exciting to see more people embracing this powerful treatment for better health and longevity
ResponElimina